Рис.
2. Показатели активности вегетативной
нервной системы.
В) Спектральная
мощность ЭЭГ.
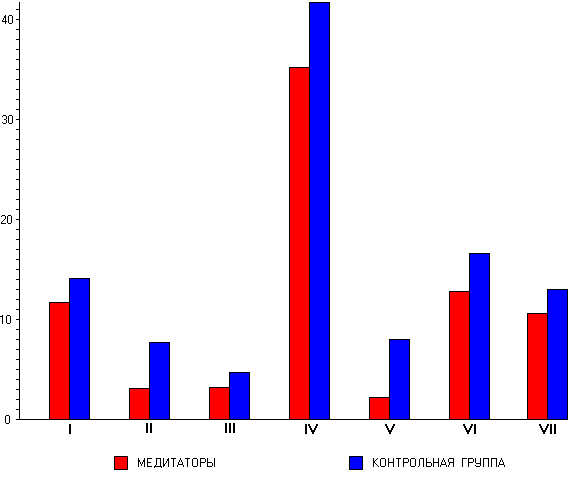
Только в состоянии покоя с закрытыми
глазами группа медитаторов по сравнению с
контролем характеризуется более высокими
значениями мощности в тета-1- (F(1,55)=5,549, p<0,002),
тета-2- (F(1,55)=5,063, p<0,028) и альфа-1- (F(1,55)=5,495, p<0,023)
диапазонах. При этом, в тета-1- и тета-2-
диапазонах преобладание мощности
наблюдается в передне-центральных, а в
альфа-1-диапазоне - во всех корковых
областях. В то же время в гамма-диапазоне
как при закрытых, так и открытых глазах у
медитаторов наблюдаются более низкие
значения мощности во всех областях коры (F(1,55)=4,578,
p<0,037).
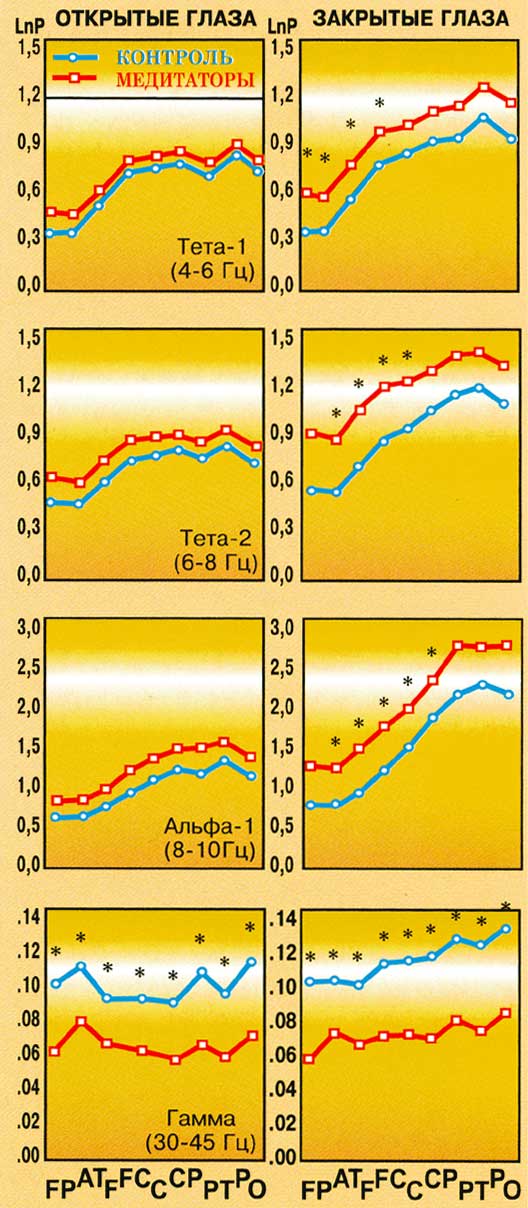
Рис. 3.
Топографические особенности распределения
мощности ЭЭГ в тета-1-, тета-2-, альфа-1- и гамма-диапазонах
у контрольных испытуемых и медитаторов в
состоянии покоя с закрытыми и открытыми
глазами. По вертикали - логарифм значения
мощности (lnP), по горизонтали - области коры (FP,
AT, F, FC, C, CP, PT, P, O). * - достоверные межгрупповые
различия по данным плановых сравнений при p<0,01
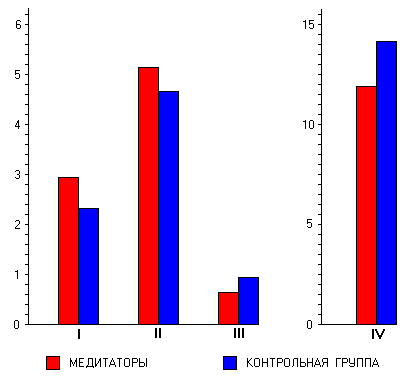
Таким образом, по данным
психометрии медитаторы по сравнению с
контрольными испытуемыми, обладая большей
интровертированностью, характеризуются
более высокой психоэмоциональной
стабильностью и лучшими способностями к
идентификации и выражению эмоций. На
вегетативном уровне по данным анализа
частоты сердечных сокращений медитаторы
характеризуются меньшими значениями
симпатической активации и более выраженной
вариативностью сердечного ритма. Более
низкая амплитуда спонтанных реакций ЭКП
также указывает на меньшую тоническую
активность симпатической нервной системы.
По данным ЭЭГ медитаторы демонстрируют
более высокие значения тета-1- и тета-2-мощности
в передних областях коры. В альфа-дипазоне
медитаторы характеризуются более высокими,
а в гамма-диапазоне более низкими
значениями мощности во всех областях коры.
2)
Спектры мощности, когерентность и
фрактальная размерность ЭЭГ в состоянии
медитации.
А) Субъективный отчёт.
Согласно данным субъективных оценок
медитаторы достигали значимо большей, чем
контроль, интенсивности переживания
состояния "блаженства" (5,54 vs. 3,56, p<0,014)
и более низкого уровня мыслительной
активности (1,19 vs. 2,28, p<0,025).
Б)
Спектральная мощность ЭЭГ.
При сравнении состояния покоя и медитации
для симметричных корковых зон левого и
правого полушария было обнаружено значимое
взаимодействие факторов ГРхЭУ в тета-2- (F(1,25)=5,299,
p<0,030) и альфа-1- (F(1,25)=5,923,
p<0,022) диапазонах, а так же ГРхЭУхЛОК в
альфа-2-диапазоне (F(5,125)=5,914,
p<0,010). У медитаторов наблюдался рост
мощности в тета-2- и альфа-1- диапазонах
преимущественно в передних отделах коры, а
в альфа-2-диапазоне - в задних отделах коры.
Контрольные испытуемые демонстрировали
снижение мощности в альфа-1- и в альфа-2-
диапазонах в задних отделах коры. При
анализе значений спектральной мощности в
электродных кластерах средней линии также
было обнаружено взаимодействие ГРхЭУ в
тета-2- (F(1,25)=5,266,
p<0,030), альфа-1- (F(1,25)=4,526,
p<0,043) диапазонах и ГРхЭУхЛОК в тета-1- (F(5,125)=4,112,
p<0,029), альфа-2- (F(5,125)=3,759,
p<0,030) диапазонах. У медитаторов наблюдался
рост тета-1-, тета-2- и альфа-1-мощности в
передне-медиальных зонах, а альфа-2-мощности
в затылочно-медиальной зоне. В результате
корреляционных анализов были обнаружены
значимые связи между изменениями
спектральной мощности в тета-1-, тета-2-,
альфа-1- и бета-1-диапазонах и оценками
субъективных характеристик медитации.
Интенсивность положительных эмоциональных
переживаний в условиях медитации
положительно коррелирует с ростом тета-1-мощности
в передних и центральных корковых зонах (значимые
корреляции в диапазоне от r=+0,38 до r=+0,41). В
свою очередь, интенсивность мыслительной
активности отрицательно коррелирует с
изменениями тета-2-мощности (значимые
корреляции от r=-0,38 до r=-0,52) по всему
корковому плану и альфа-1- (от r=-0,38 до r=-0,52),
бета-1-мощностью (от r=-0,45 до r=-0,49) в передних
отведениях.
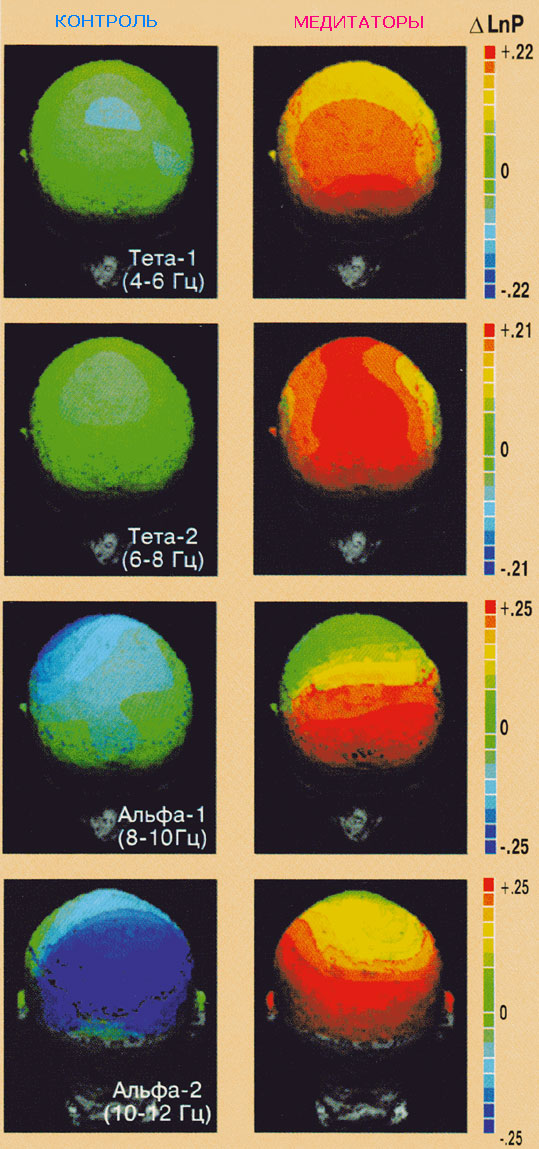
Рис. 4.
Изменение мощности ЭЭГ в тета-1-, тета-2-,
альфа-1- и альфа-2-диапазонах между
состоянием покоя с закрытыми глазами у
контрольных испытуемых и состоянием
медитации у медитаторов.
В) Когерентность ЭЭГ.
Эффекты медитации были обнаружены только в
тета-1-диапазоне (4-6 Гц). В условиях медитации
группа медитаторов характеризовалась
усилением коротко- и длиннодистантных
когерентных связей преимущественно в
передне-заднем направлении с выраженным
"центром тяжести" левой
префронтальной области.
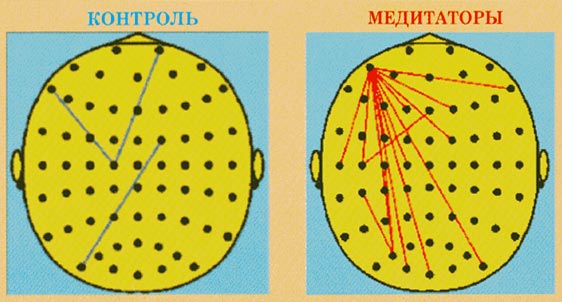
Рис.5.
Изменения когерентности в тета-1-диапазоне
(4-6 Гц) между состояниями покоя и медитации
для групп контроля и медитаторов. Красные
линии указывают увеличение когерентности,
синие - уменьшение когерентности при p<0,01
Г)
Фрактальная размерность ЭЭГ.
В условии медитации медитаторы
демонстрируют фокусированное снижение
оценок DCx в передних отведениях средней
линии, что подтверждается значимым
взаимодействием ГРхЭУ (F(1,25)=4,982,
p<0,036) и результатами последующего
внутригруппового анализа для группы
медитаторов (F(5,75)=5,400,
p<0,012). У медитаторов оценки DCx и значения
мощности средней линии отрицательно
коррелировали между собой в тета-2- (от r=-0,52 до r=-0,57)
и альфа-1- (от r=-0,56 до r=-0,66) диапазонах. Таким
образом, состояние медитации
характеризуется усилением тета-1-, тета-2- и
альфа-1-мощности в передних областях коры,
наряду со снижением оценок фрактальной
размерности, а в задних областях коры -
ростом альфа-2-мощности. Рост тета-1-мощности
положительно коррелирует с выраженностью
положительных эмоциональных переживаний, а
изменения тета-2-, альфа-1- и бета-1-мощности
отрицательно коррелирует с интенсивностью
мыслительной активности. Кроме того,
состояние медитации сопровождается ростом
длиннодистантной тета-1-когерентности
между префронтальной и задней
ассоциативной корой с формированием "центра
тяжести" в левой префронтальной области.
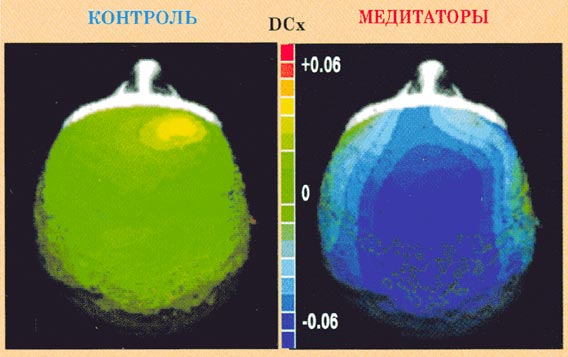
Рис.6.
Изменения оценок фрактальной размерности (DCx)
между состоянием покоя с закрытыми глазами
и медитации у контрольных испытуемых и
медитаторов.
3)
Вызванная синхронизация и десинхронизация
в процессе опознания эмоциональности
сигнала.
А) Субъективный отчёт
В оценке знака (F(2,80)=18,479,
p<0,001) и активационного содержания (F(2,80)=18,479,
p<0,001) стимулов обнаруживаются отчётливые
межгрупповые различия - медитаторы по
сравнению с контрольными испытуемыми
оценивают положительные и отрицательные
стимулы как менее эмоциогенные.
Б)
Данные вызванной синхронизации и
десинхронизации.
Вне зависимости от категории стимула,
медитаторы характеризуются меньшими
значениями ВС в дельта- (F(1,39)=11,183, p<0,002),
тета-1- (F(1,39)=4,184, p<0,048), тета-2- (F(1,39)=11,577,
p<0,002) и гамма- (F(1,39)=9,667,
p<0,003) диапазонах. В тета-1- и гамма-диапазонах
медитаторы и контрольные испытуемые
обнаружили сходную топографическую и
временную структуру реакции ВС на
эмоциогенные стимулы: эффекты
эмоциогенности стимула проявляются
преимущественно в задних областях коры в
период 0-700 мс. Однако, в дельта-диапазоне в
теменно-височных-затылочных областях коры
обнаружены специфические межгрупповые
различия, связанные с эмоциогенностью
стимула и функциональной полушарной
асимметрией (F(12,468)=3,803,
p<0,009). В ответ на положительные и
отрицательные стимулы медитаторы
демонстрируют меньшие значения ВС
билатерально по всему временному интервалу.
Наряду с этим, наблюдаемая у контрольных
испытуемых более выраженная синхронизация
в правом полушарии в ответ на негативные
эмоциогенные стимулы в интервале 400-700 мс. от
начала предъявления отсутствует у
медитаторов.
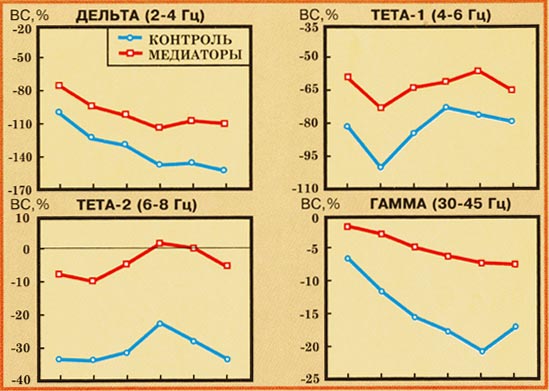
Рис. 7.
Особенности топографического
распределения эффектов ВС в дельта-, тета-1-,
тета-2- и гамма-диапазонах у медитаторов и
контроля в ответ на предъявление стимулов
вне зависимости от эмоциональной категории
в тестовом интервале 0-700 мс. По горизонтали -
области коры (AT, F, C, PT, P, O); по вертикали - ВС (%,
усиление синхронизации вниз от нулевой
линии). * - достоверные различия по данным
плановых сравнений при p<0,01
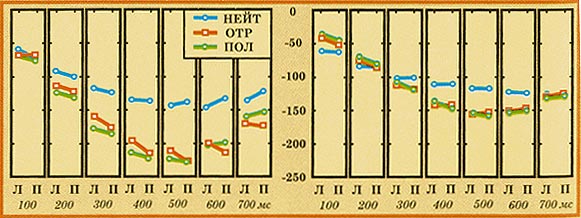
Рис.
8. Временная динамика ВС в задних областях
левого (Л) и правого (П) полушарий
мозга в ответ на предъявление нейтральных (НЕЙТ),
отрицательных (ОТР) и положительных (ПОЛ)
стимулов в дельта-диапазоне (2-4 Гц) в
интервале 0-700 мс для групп медитаторов и
контроля. По горизонтали - время от начала
предъявления стимула; по вертикали ВС (%,
усиление синхронизации вниз от нулевой
линии).
Таким образом, на
субъективном уровне эмоциогенные
сигналы оказывают меньшее активационное
воздействие на медитаторов по сравнению с
контрольными испытуемыми. Процесс
эмоционального восприятия у медитаторов
характеризуется более "экономным"
использованием активационных ресурсов: вне
зависимости от категории стимула,
медитаторы демонстрируют более низкие
значения синхронизации в дельта-, тета-1-,
тета-2- и гамма-диапазонах. В дельта-диапазоне
в теменно-височно-затылочных областях коры
билатерально у медитаторов наблюдаются
более низкие значения связанной с
эмоциогенностью вызванной синхронизации.
Кроме того, характерная для контрольных
испытуемых более выраженная синхронизация
в правом полушарии в ответ на негативные
эмоциогенные стимулы, отсутствует у
медитаторов.
4) Спектры мощности
ЭЭГ в модели переживания дискретных эмоций.
А) Субъективный отчёт.
До начала стимуляции значимых различий в
эмоциональном профиле контрольных
испытуемых и медитаторов обнаружено не
было. Внутригрупповые анализы подтвердили
наличие роста значений "целевых"
эмоций (например, радость при индукции
радости) при предъявлении эмоциональных
клипов по сравнению с нейтральными в обеих
группах испытуемых. Однако, на межгрупповом
уровне медитаторы демонстрировали значимо
менее выраженную интенсивность
переживания эмоций веселья (F(9,486)=4,51,
p<0,001), гнева (F(9,486)=6,28,
p<0,001), отвращения (F(9,486)=2,70,
p<0,005) и грусти (F(9,486)=3,80,
p<0,001).
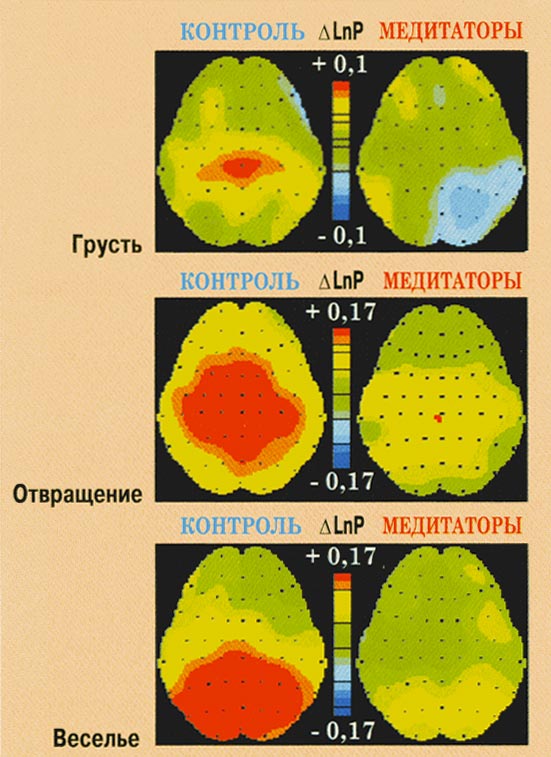
Рис. 9. Изменения мощности ЭЭГ в
тета-2-диапазоне при переживании эмоций
веселья, отвращения и грусти у контрольных
испытуемых и медитаторов.
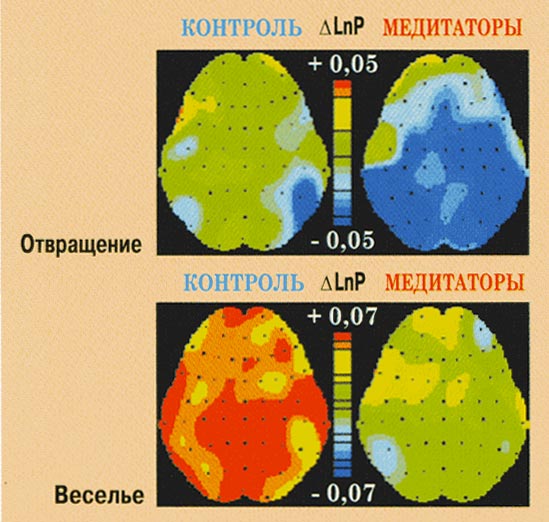
Рис. 10. Изменения мощности ЭЭГ в
бета-1-диапазоне при переживании эмоций
веселья и отвращения у контрольных
испытуемых и медитаторов.
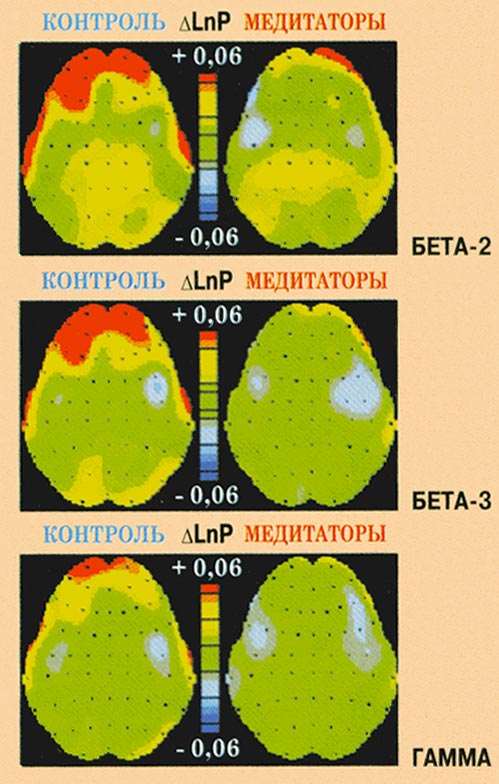
Рис. 11. Изменения мощности ЭЭГ в
бета-2-, бета-3- и гамма-диапазонах при
переживании эмоции гнева у контрольных
испытуемых и медитаторов.
Б) Спектры мощности ЭЭГ.
Эмоция веселья.
Переживание эмоции веселья вне зависимости
от фактора группы сопровождалось ростом
тета-1- (F(1,55)=10,760,
p<0,002) и бета-2- (F(1,55)=5,610,
p<0,021) мощности по всему корковому плану, а
также ростом бета-3- (F(4,220)=4,400,
p<0,002) и гамма- (F(2,110)=5,350,
p<0,006) мощности в центральных и задних
отделах коры. Кроме того, у испытуемых обеих
групп в альфа-1-диапазоне наблюдалось
снижение мощности в центральных и задних
областях (F(4,220)=9,250,
p<0,001). Анализ взаимодействия ГРхЭУхОК (F(2,110)=5,997,
p<0,008) в тета-2-диапазоне указывает на рост
мощности в задних областях коры у
контрольных испытуемых и отсутствие
значимых изменений у медитаторов. Напротив,
в альфа-2-диапазоне взаимодействие ГРхЭУ (F(1,55)=8,520,
p<0,005) обусловлено снижением мощности у
медитаторов и отсутствием изменений
мощности в группе контроля. В бета-1-диапазоне
взаимодействие ГРхЭУ (F(1,55)=5,352,
p<0,024), как и в случае тета-2-полосы,
обеспечивается реактивностью контрольных
испытуемых и ареактивностью медитаторов.
В)
Эмоция гнева.
В обеих группах переживание эмоции гнева
сопровождалось билатеральным усилением
тета-2-мощности (F(2,110)=12,130,
p<0,001) в центральных и задних областях коры.
В бета-1-диапазоне наблюдался асимметричный
рост мощности в левом полушарии (F(1,55)=5,897,
p<0,018). Характерной особенностью
медитаторов явилось отсутствие
наблюдаемого у контрольных испытуемых
роста бета-2- (F(2,110)=4,213,
p<0,029), бета-3- (F(2,110)=6,814,
p<0,002) и гамма- (F(2,110)=5,218,
p<0,007) мощности в передних областях коры.
Г)
Эмоция отвращения.
У обеих групп эмоция отвращения
характеризовалась эффектами альфа-1- (F(2,110)=21,003,
p<0,001) и альфа-2- (F(2,110)=8,550,
p<0,001) десинхронизации в центральных и
задних областях коры. В тета-2-диапазоне (F(1,55)=5,346,
p<0,025) медитаторы не обнаруживают значимых
изменений мощности, в то время как у
контроля мощность растёт по всему
корковому плану. В бета-1-диапазоне (F(2,110)=3,392,
p<0,041) в задних областях коры у медитаторов
наблюдается снижение мощности, а у
контрольных - отсутствие реакции.
Д)
Эмоция грусти.
Эмоция грусти характеризовалась у обеих
групп испытуемых усилением альфа-2- (F(2,110)=4,327,
p<0,026) и бета-1- (F(4,220)=9,331,
p<0,001) мощности преимущественно в задних
отведениях коры. В тета-2-диапазоне (F(1,55)=8,175,
p<0,006) у медитаторов отсутствует
наблюдаемый у контроля рост мощности. В
отличии от контроля медитаторы
демонстрируют снижение альфа-1-мощности (F(2,110)=4,163,
p<0,018) в задних областях коры. В
высокочастотной области спектра (бета-3- F(1,55)=4,414,
p<0,041; гамма - F(2,110)=3,660,
p<0,045) только контрольные испытуемые
демонстрируют значимые изменения -
десинхронизацию в центрально-теменных
областях коры. Таким образом, на
субъективном уровне медитаторы
демонстрируют меньшую интенсивность
переживания вызванных дискретных эмоций.
По данным ЭЭГ, переживание вызванных
положительных и отрицательных эмоций
медитаторами по сравнению с контролем
характеризуется:
-
отсутствием эффектов усиления
тета-2-мощности в задних (эмоция веселья)
либо во всех (эмоция грусти и отвращения)
областях коры;
-
снижением бета-1-мощности в
задних (эмоция отвращения) и отсутствием
её усиления (эмоция веселья) в передних и
задних корковых областях;
-
отсутствием роста бета-3- и
гамма-мощности (эмоция гнева) в передних
областях коры головного мозга и снижения
(эмоция грусти) - в центрально-теменных
областях.
5) Спектры мощности ЭЭГ и
вегетативная реактивность (ЭКП) в условиях
стрессовой стимуляции.
А) Субъективный отчёт.
На субъективном уровне, в условиях
стрессовой стимуляции у обеих групп
испытуемых наблюдался значимый рост
интенсивности переживаний отрицательных
эмоций (гнева, страха, тревоги, отвращения,
отторжения), однако, у медитаторов
амплитуда реакций была достоверно ниже (F(9,477)=12,44,
p<0,001)
Б) Спектры мощности ЭЭГ.
Стрессовая стимуляция в обеих группах
приводила к альфа-1- (F(2,106)=10,748,
p<0,001), альфа-2- (F(2,106)=6,188,
p<0,004) десинхронизации в центрально-теменных
и гамма- (F(2,106)=4,508,
p<0,027) синхронизации в передне-центральных
областях коры. В бета-1-диапазоне (F(1,53)=5,237,
p<0,026) медитаторы обнаруживают выраженную
десинхронизацию, в то время как у контроля
значимые изменения в этой частотной полосе
отсутствуют.
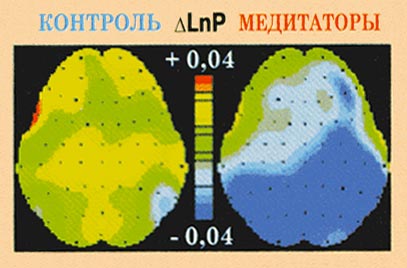
Рис. 12. Изменения мощности ЭЭГ в
бета-1-диапазоне в ответ на предъявление
стрессирующего видеоклипа по отношению к
нейтральному у контрольных испытуемых.
В) Электрокожный
потенциал.
На вегетативном уровне стрессовая
стимуляция приводила к достоверно более
выраженному росту амплитуды уровня
электрокожного потенциала у контрольных
испытуемых по сравнению с медитаторами
(p<0,01). А по данным внутригрупповых
анализов, изменения электрокожного
потенциала в ответ на стрессовый вызов по
сравнению с условием нейтральной
стимуляции были значимыми только в группе
контроля (p<0,04). Помимо этого были
обнаружены положительные корреляции между
изменением спектральной бета-1-мощности ЭЭГ
и амплитудой электрокожных реакций (диапазон
от r=0,27 до r=0,42) в условиях стрессовой
стимуляции. Таким образом, по данным
субъективных оценок в условиях
психоэмоционального стресса медитаторы
характеризовались более низкой
интенсивностью переживаний эмоций
отрицательного круга. Стрессозащитные
эффекты медитации на вегетативном уровне
подтверждаются более низкими уровнями
симпатической активации по данным
электрокожной активности, а на корковом
отражаются в снижении активности ЭЭГ в бета-1-диапазоне.
Результаты корреляционного анализа
подтверждают связь между изменениями
мощности ЭЭГ в бета-диапазоне и амплитуды
ЭКП: более выраженное снижение мощности ЭЭГ
ассоциировалось с минимизацией реакций ЭКП
на стрессовый вызов.
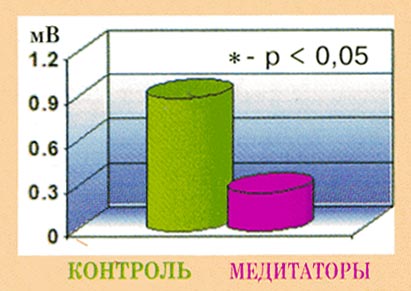
Рис. 13.
Изменения амплитуды реакций
электрокожного потенциала (ЭКП) в ответ на
предъявление стрессирующего видеоклипа по
отношению к нейтральному у контрольных
испытуемых.
ВЫВОДЫ
-
По сравнению с контрольными
испытуемыми на личностном уровне
медитаторы (медитация по системе Сахаджа
Йога), обладая большей
интровертированностью, отличаются более
высокой психоэмоциональной
стабильностью и лучшими способностями к
идентификации и выражению эмоций.
-
В состоянии физиологического
покоя по данным ЭЭГ медитаторы
демонстрируют более высокие значения
тета-1- и тета-2-мощности в передних, а
альфа-1-мощности - во всех областях коры. В
диапазоне гамма-активности наблюдается
общее снижение мощности по всему
корковому плану. По данным значений
электрокожного потенциала и
вариативности сердечного ритма
медитаторы обнаруживают сниженные
уровни вегетативной активации.
-
Состояние медитации
характеризуется усилением спектральной
мощности в тета-1-, тета-2- и альфа-1-диапазонах
наряду с фокусированным снижением оценок
фрактальной размерности ЭЭГ в передне-медиальных
областях коры, а также ростом
длиннодистантной тета-когерентности
между передней и задней ассоциативной
корой с формированием "центра тяжести"
в левой префронтальной области.
-
Субъективные показатели
положительных эмоциональных переживаний
в состоянии медитации положительно
коррелируют с изменением тета-1-мощности,
а торможение когнитивной активности - с
изменением тета-2-мощности в передне-медиальных
областях коры.
-
Процесс восприятия у
медитаторов отличается более "экономным"
использованием активационных ресурсов.
-
Межгрупповые различия
эмоционального восприятия
обнаруживаются в диапазоне дельта-активности
в теменно-височно-затылочных областях
коры: в ответ на положительные и
отрицательные эмоциональные стимулы по
сравнению с нейтральными медитаторы
демонстрируют меньшие значения
вызванной синхронизации. Наряду с этим, у
медитаторов отсутствует наблюдаемая у
контрольных испытуемых более выраженная
синхронизация в правом полушарии в ответ
на негативные эмоциогенные стимулы.
-
На субъективном уровне
медитаторы демонстрируют меньшую
интенсивность переживания вызванных
дискретных эмоций. Нейрофизиологически,
особенности реализации эмоций
отражаются в изменениях реактивности в
тета-2-, бета-1-, бета-3- и гамма-диапазонах.
-
В условиях психоэмоциональной
стрессовой стимуляции защитные эффекты
медитации на вегетативном уровне
подтверждаются более низкими уровнями
симпатической активации по данным
электрокожной активности, а на корковом -
отражаются в снижении мощности ЭЭГ в бета-диапазоне.
Список сокращений:
ВД/ВС - вызванная десинхронизация/синхронизация;
ВСР - вариативность сердечного ритма;
ЭКГ - электрокардиограмма;
ЭКП - электрокожный потенциал;
ЭЭГ - электроэнцефалограмма;
DCx - фрактальная размерность.